Cartilage Histology
Summary of Cartilage Types & Features:
Hyaline Cartilage: composed of type II collagen, basophilic matrix, arranged in isogenous groups; found at the articular end of long bones in joint capsules, larynx, trachea; Perichondrium present in most places (Except articular ends of long bones)
Elastic Cartilage: composed of type II collagen, elastic fibers; found in ear pinna, auditory tube, epiglottis; Perichondrium is Present
Fibrocartilage: composed of type I collagen, acidophilic matrix, associated with dense regular connective tissue. Chondrocytes are arranged in parallel rows; Perichondrium is Absent
I. Cartilage Structure & Associated Cells
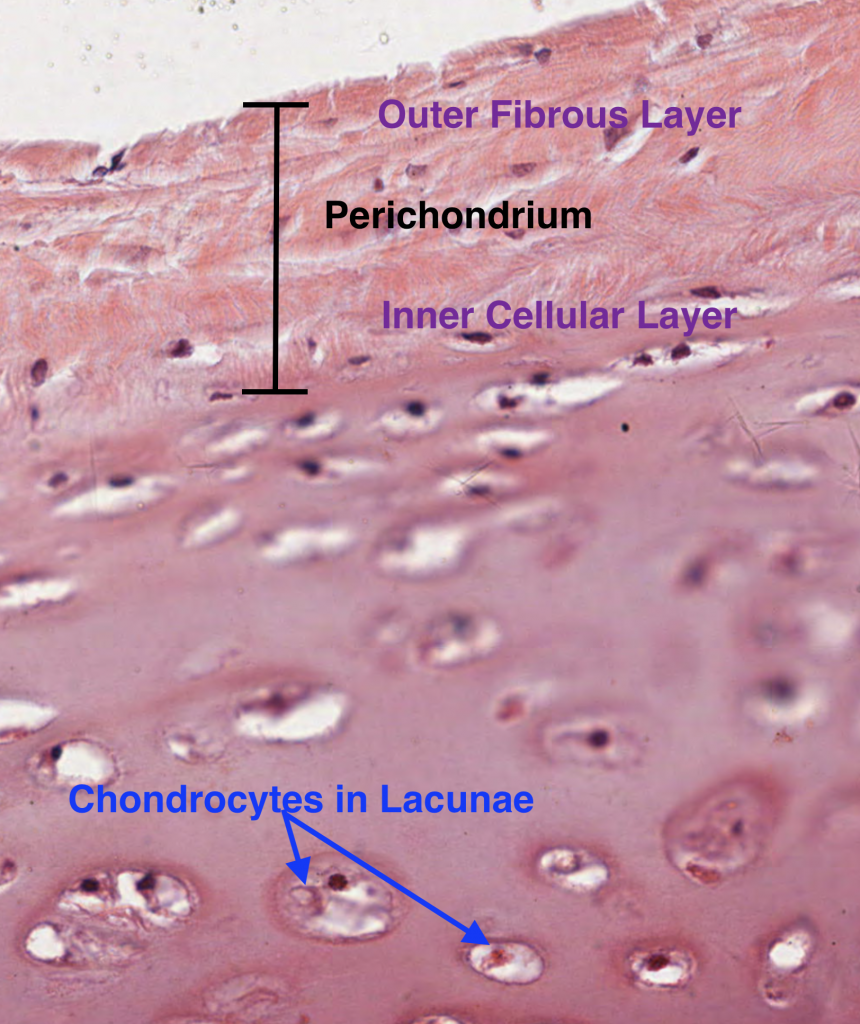
Cartilage consists of chondrocytes, which are cells that secrete an extracellular matrix (proteoglycans and glycosaminoglycans) that eventually surrounds themselves into small chambers called lacunae. Cartilage is an avascular connective tissue and requires nourishment of the chondrocytes within by diffusion of nutrients from the blood vessels found in perichondrium. The perichondrium is a dense irregular connective tissue sheath overlying most cartilage found in the body. The only place where a perichondrium does not exist is within joint capsules (ex: joints of the ankle, knee, hip and shoulder). Here, the chondrocytes must be nourished by synovial fluid secreted by synovial membranes lining the joint capsules.
The perichondrium can be subdivided into two layers: outer fibrous layer and inner cellular layer.
Outer fibrous layer: type I collagen, fibroblasts, and blood vessels
Inner cellular layer: chondrogenic cells are derived from mesenchymal cells and are destined to become chondrocytes. They are spindle-shaped and appear narrow compared to the mature chondrocytes. The chrondrogenic cells of the inner cellular layer are stacked above the chondrocytes below and eventually develop into matrix-secreting chondrocytes. Therefore, new cartilage matrix is deposited from the outside in, which is known as Appositional Growth.
In the absence of perichondrium, growth occurs by interstitial growth, in which, chondrocytes are aligned longitudinally along the long axis of a bone and secrete matrix such that the bone lengthens. This type of growth is typical of epiphyseal plates of long bones.
Hyaline Cartilage:
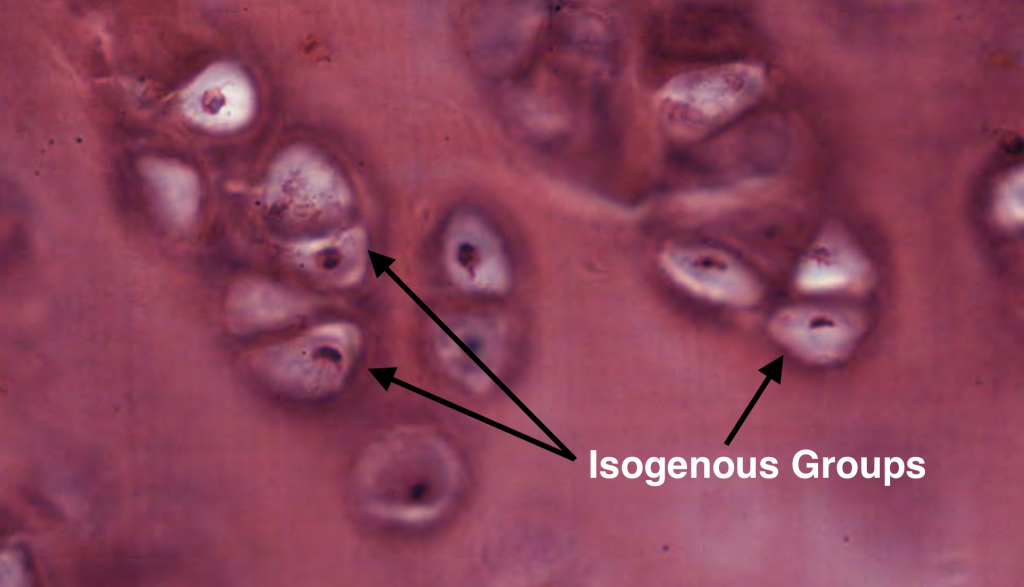
Isogenous groups represent a single chondrocyte that has undergone several rounds of cell division depending on how many chondrocytes are found within each group. Therefore, each chondrocyte of the isogenous group are genetically identical. Each chondrocyte within the isogenous group will eventually secrete its own matrix and break away from the group. This contributes to the interstitial growth of long bones.
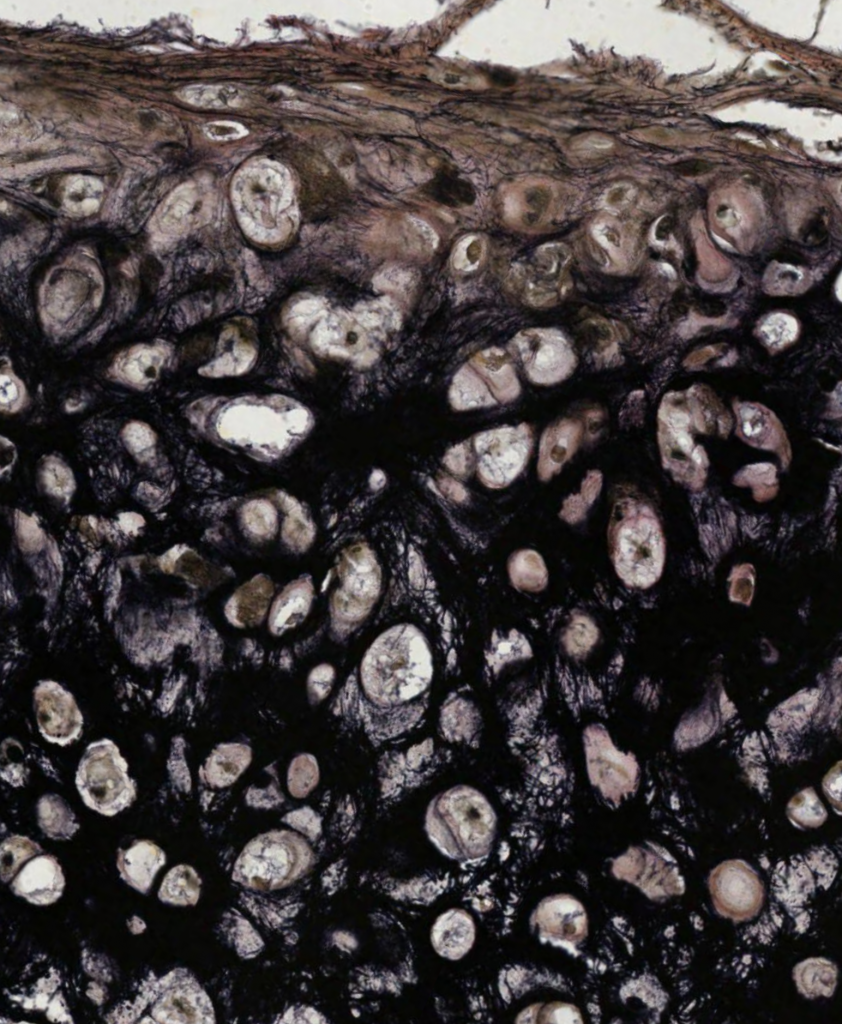
Elastic Cartilage with Elastin Stain:
Fibrocartilage:
Chondrocytes can be seen with a pale matrix relative to the eosinophilic staining of the type I collagen fibers found throughout the tissue section. Fibrocartilage is typically found in the intervertebral disc, which is depicted in the specimen below.
Bone Histology
Bone consists of an outer layer of periosteum (subdivided into an outer fibrous & inner cellular layer), similar to cartilage, but the inner cellular layer consists of osteoprogenitor cells instead chondrogenic cells. Articular bone regions covered by hyaline cartilage do not have a periosteum. The periosteum is anchored to the bone by special bundles of collagen fibers known as Sharpey’s fibers. The inner medullary cavity of bone is lined with endosteum composed of a single layer of osteoprogenitor cells and osteoblasts (bone matrix-secreting cells).
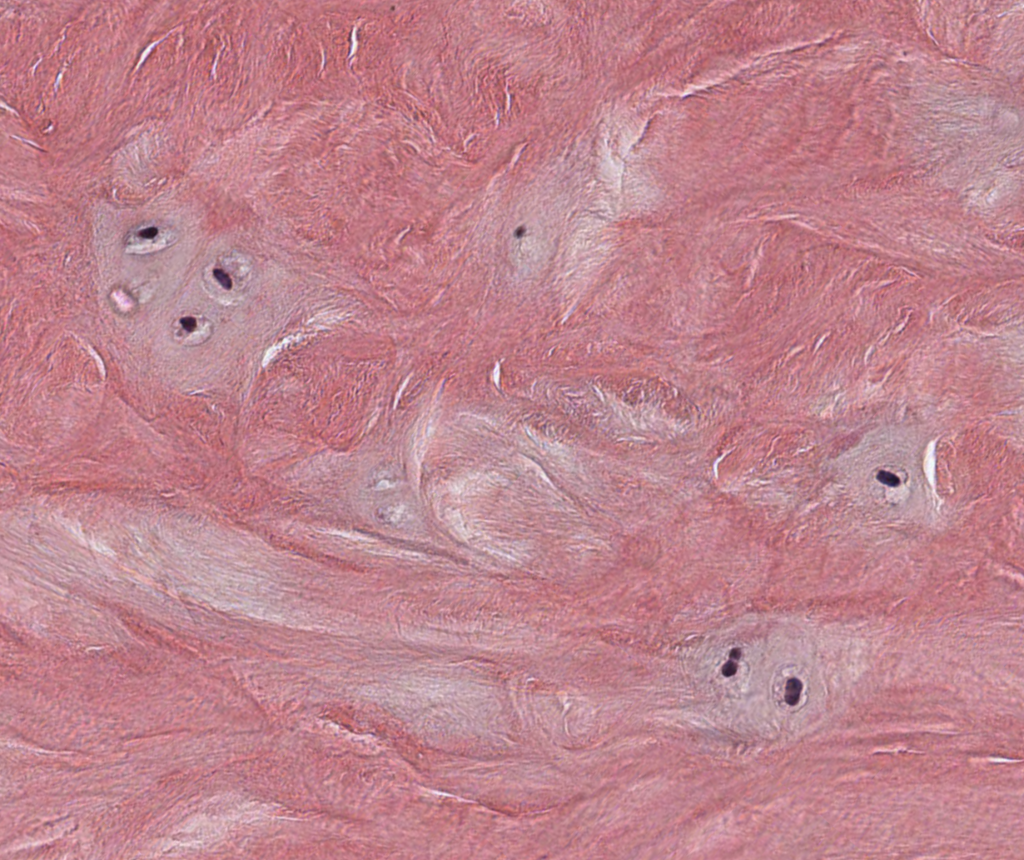
I. Compact Bone
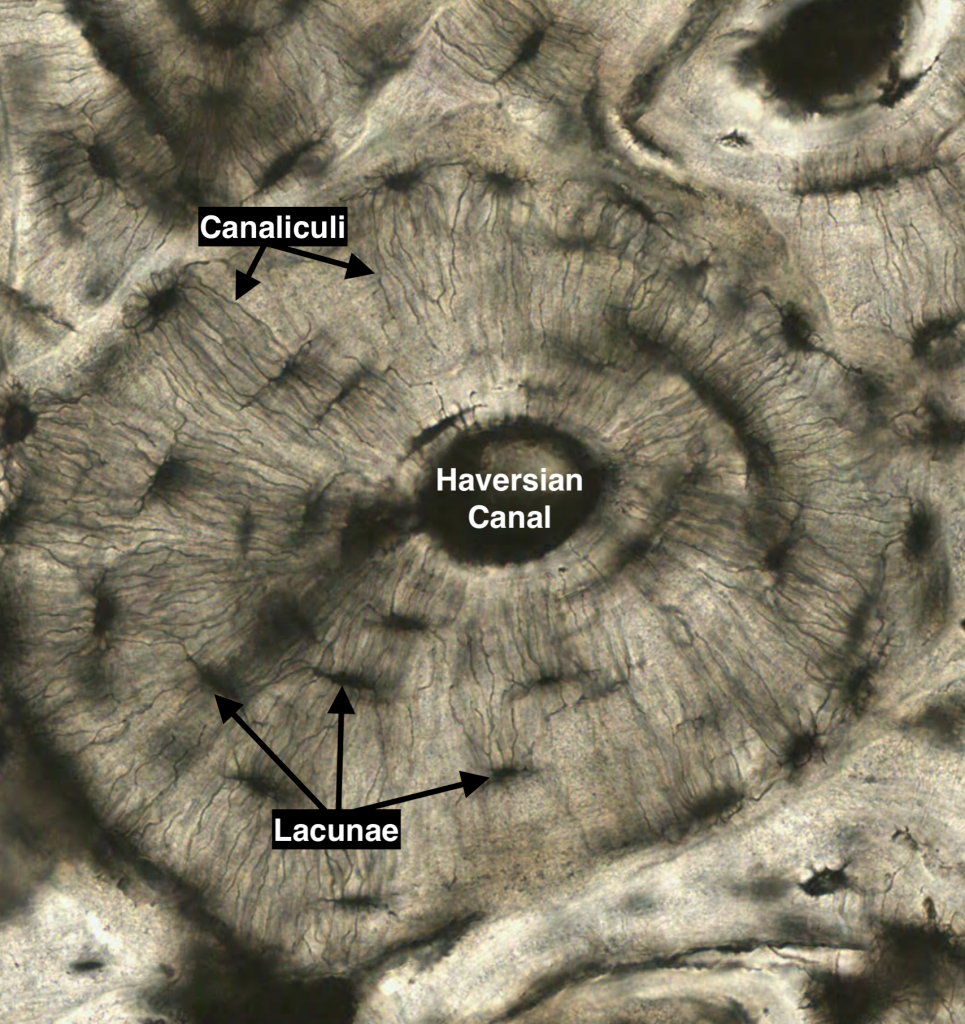
Compact bone consists of groups of lamellar cylinders (osteons) that contain a central vascular space known as the Haverisian Canal. Adjacent blood vessels are connected through oblique or perpendicularly-arranged channels known as Volkmann’s canals. The osteocytes secrete matrix and become encapsulated in the their lacunae, and they connect with adjacent lacunae through cytoplasmic extensions known as canaliculi. Nutrients must be passed from the blood vessels of the central Haversian canal through canaculi and subjacent lacunae to reach all the osteocytes found in the outer lamellar regions.
II. Spongy Bone
Please Histogenesis of Bone for further detail in the laboratory manual
Skeletal Muscle Histology
I. Muscle Structure
Skeletal muscle fibers are fusiform (spindle-shaped) structures consisting of aggregations of several contractile cells that have fused together during embryological development. Specifically, the skeletal muscle precursor cells, myoblasts, align and fuse together to form multinucleated myotubes. The myotubes continue to develop cytoplasmic structures and the contractile elements known as myofibrils.
Each myofibril is composed of discrete contractile units –sarcomeres – which are arranged in series. The striated muscle appearance is due to overlapping and non-overlapping actin and myosin filaments. Muscles insert on bone and allow for movement of joints through contractions produced by the cumulative shortening output of the sarcomeres of the myofilaments found within each muscle fiber. The sarcomeres can be appreciated by looking at an electron micrograph of skeletal muscle, in which, the overlapping actin and myosin filaments can be seen.

Image: Gartner & Hiatt (2007). Color Textbook of Histology: Saunders Elsevier, p. 464

Skeletal muscles fibers contain striations and peripherally-located nuclei.
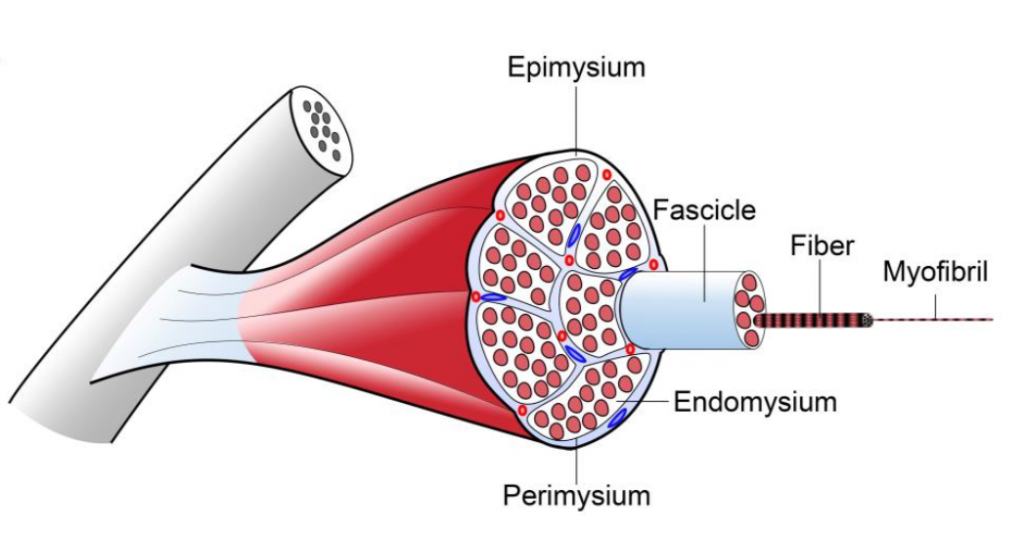
II. Connective Tissue Sheaths
Skeletal muscle is an organized tissue with distinct connective tissue sheaths at several levels of skeletal muscle structure known as the epimysium, perimysium and endomysium.
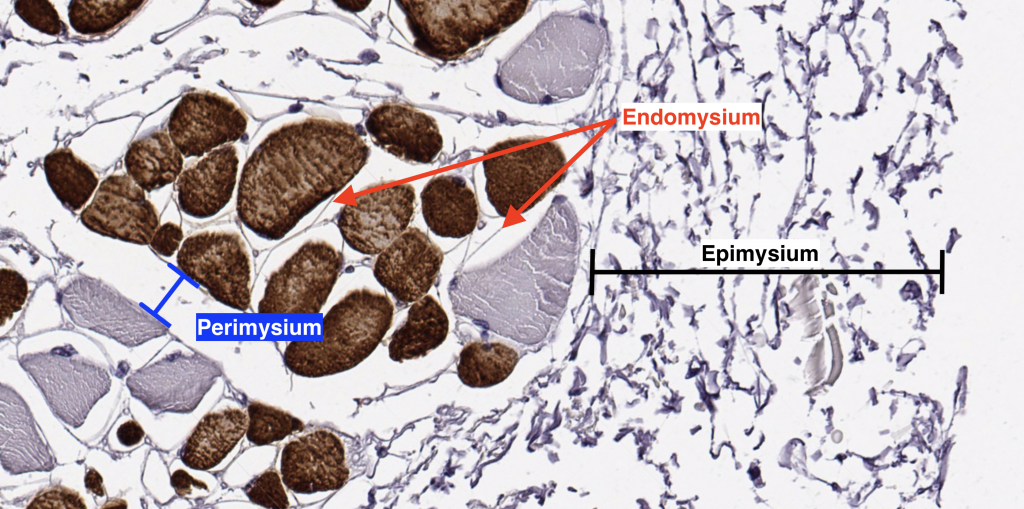
Epimysium: a thick layer of dense connective tissue that surrounds the whole muscle and becomes continuous with the tendon
Perimysium: a thick connective tissue layer that surrounds muscle fascicles; larger blood vessels and nerves travel within the perimysium
Endomysium: a delicate layer of reticular fibers that surrounds the individual muscle fibers within a muscle fascicle. Small neuronal fibers and blood vessels may be found in this layer.
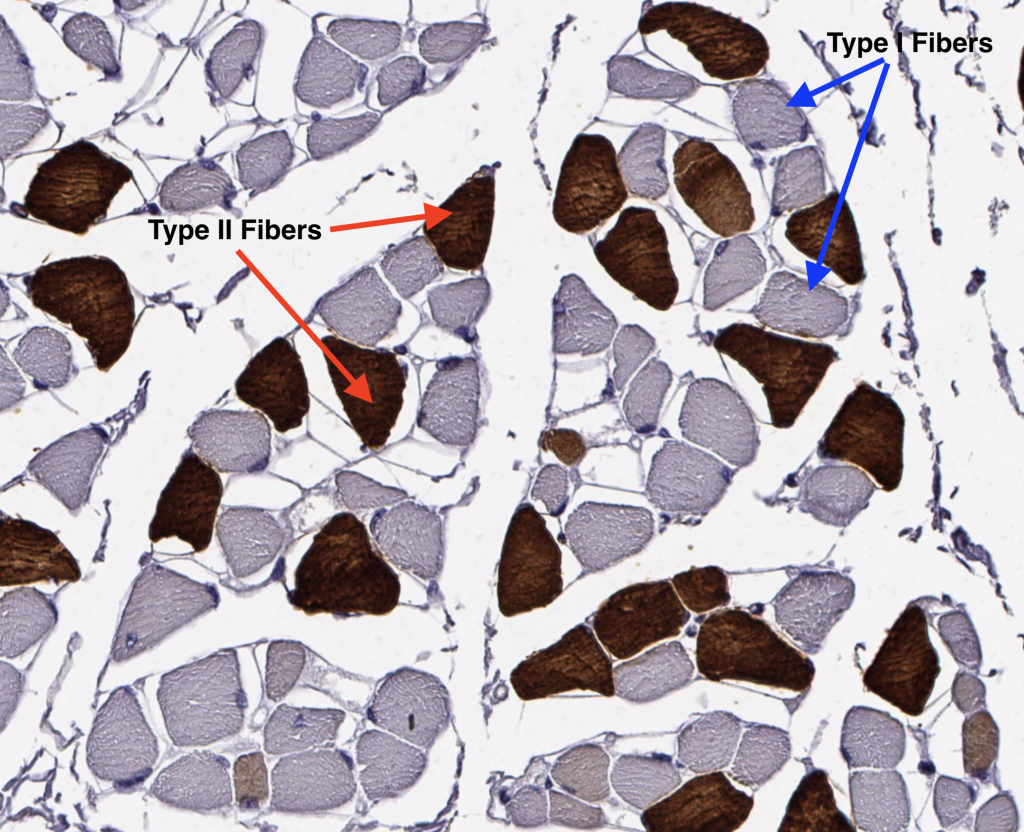
III. Muscle Fiber Types
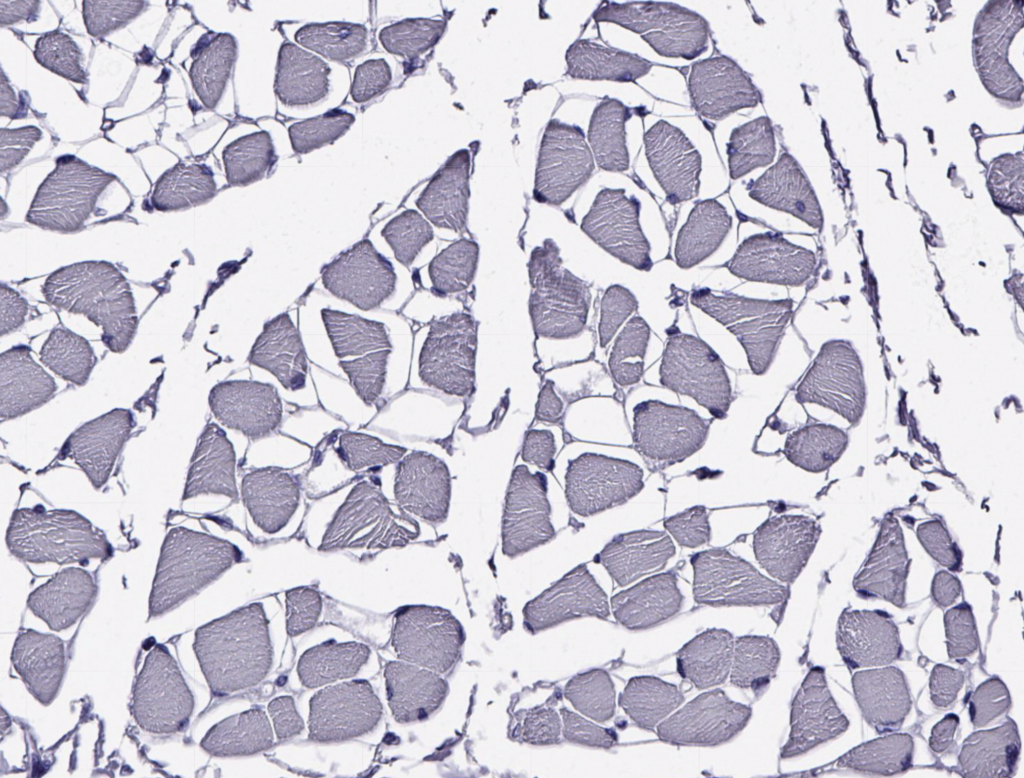
A skeletal muscle typically contains three types of muscle fibers each with specific metabolic, structural and contractile properties. Different muscles contain these three fiber types but in varying proportions based on the functional demands placed on the muscle. Orbicularis oculi muscles contain >80% fast twitch (Type II) muscle fibers for rapid closing of the eye. Conversely, the soleus muscle, which is a muscle of postural stability and constantly active in gait, consists of >80% slow twitch (Type I) muscle fibers. The fiber types cannot be appreciated without staining for the constituent myosin heavy chain (MHC) isoforms or oxidative/glycolytic enzymes, or myosin ATPase. Below, an unstained muscle stained with hematoxylin only demonstrates that the individual fiber identities cannot be seen without adding a special stain.